























-
1
-
2
-
3
-
4
-
5
-
6
-
7
-
8
-
9
-
10
-
11
-
12
해당 자료는 4페이지 까지만 미리보기를 제공합니다.
4페이지 이후부터 다운로드 후 확인할 수 있습니다.
4페이지 이후부터 다운로드 후 확인할 수 있습니다.




목차
없음
본문내용
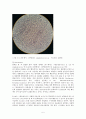
staphylococcus가 늙은 배양균이기 때문에 염색성이 좋지 않아 음성균같이 빨간색으로 보였을 수 있다. 또한 그람염색법은 숙련될 때까지 많은 연습이 필요하다고 하므로 아직 기술 부족으로 인해 염색이나 탈색을 제대로 못했기 때문인것같다. 또 빨간색 부분은 빨간색이라기보다는 색이 바래서 옅은 빨간색으로 보이는데 그 이유는, 1번 배지를 관찰하느라, 2번 배지를 염색한 후에 너무 오랫동안 공기중에 놔두었기 때문에 색이 바랜 것 같다.
<그림 4-1. 2번 배지를 전체적으로 봤을때>
분열하는 Staphylococcus aureus 세포의 투과전자현미경 사진을 보면 두꺼운 그람양성 세균의 세포벽을 볼 수 있다.
실험에서는 대장균과 황색포도상구균으로 그람양성균과 그람음성균을 구별하는 법을 배웠지만 이외의 다른 균으로는 그람양성세균은 Lactococcus lactis,Streptococcus 종, clostridium, Sarcina 종 등이 있으며 대부분 화학종속영양생물이 있고 그람음성세균으로는 시아노박테리아, 프로테어박테리아 등이 있다.
실험과정에서 몇 가지 주의할 것이 있다. 염색약은 묻으면 잘 지워지지 않으므로 장갑을 꼭 끼고 한다. 공기중에 노출되면 오염되니까 실험하는 곳을 물이나 에탄올로 세척한다. 백금이를 화염멸균하고 균을 취한다. 이때 두껍게 도말하면 관찰하기 어려우므로 백금이에 살짝 묻히듯 취한다. 또 불 근처에서 건조시킬때 빨리 건조시키기 위해 입으로 불거나 기구를 사용해서는 안된다. 침이나 먼지, 균이 들어가 오염되기 때문이다. 글라스 위에 물을 한방울 떨어뜨린 후 그 위에 균을 풀면서 지름 1~1.5cm 정도의 원모양으로 펼쳐 도말할 때 증류수는 퍼지게 하는 역할이다. 이것은 백금이로 편다. 증류수가 너무 많으면 잘 마르지 않아 그만큼 시간이 오래 걸리고 그만큼 공기 중의 다른 미생물이 실험 중인 균주에 들어가 오염될 가능성이 크다. 도말할 때 넓게 퍼지게 할수록 관찰하기 좋다. 도말 부위를 위로 한 상태에서 불꽃 중간에 슬라이드글라스를 3~4회 통과시켜서 표본을 열고정을 해야하는데 그 이유는 세척을 할 때, 세균이 씻겨 내려가는 것을 방지하기 위해서이다. 또 그람염색 중 뒷면에 증류수로 세척한 이유는 세균이 있는 앞면에 세척을 하면, 세균이 씻겨내려갈 수 있기 때문이다. 또 그람염색에서는 시약 순서대로 하는 것과 염색과 탈색 시간을 지키는 것이 아주 중요하다. 시간이 너무 짧으면 충분히 염색이 되지 않고, 또 오랫동안 염색하면 색이 너무 진해지며, 탈색시간인 10초를 넘으면, 그람양성균의 펩티도글리칸층이 아무리 두꺼워도 녹기 때문이다.
그람염색법은 미생물의 모양과 크기를 쉽게 관찰할 수 있다. Gram이 개발한 그람염색법에서 그람양성세균과 그람음성세균간의 차이점은 세포벽 구조의 차이에 기인한다. 그람음성세균의 세포벽은 다층 구조이며 아주 복잡하지만 그람양성세균의 세포벽은 단일층으로 되어 있고 더 두꺼우며 거의 모두 한 종류의 분자로 구성되어 있다. 그람양성세균과 그람음성세균에서 세포벽의 단단한 층(rigid layer)은 화학성분이 매우 유사한데 이를 펩티도글리칸이라 부른다. 그람양성세균의 세포벽은 세포질막 바쪽으로 20~80nm두께의 펩티도글리칸 단일층으로 이루어져 있다. 펩티도글리칸의 긴 사슬구조는 세포 주위를 둘러는 판을 형성하기 위하여 하나씩 연속적으로 생합성된다. 펩티도글리칸 사슬은 아미노산의 교차결합(cross-link)에 의하여 연결된다. 글리칸 사슬에서 당을 연결하는 배당결합(glycosidic bond)은 공유결합이지만 한 방향으로만 구조의 견고성을 줄 수 있다. 펩티도글리칸의 교차결합 후에만 X 및 Y방향으로 견고하게 된다. 교차결합은 세균의 종류에 라 그 정도가 다른데 교차결합이 많아질수록 더욱 견고해진다. 원핵세포는 표층구조의 차이에 따라 4군으로 나눌 수 있다. 즉 세포벽이 펩티도글리칸 단일층으로 이루어진 그람양성세균, 세포벽이 펩티도글리칸층과 그 바깥쪽을 둘러싸고 있는 외막(outer membrane)으로 이루어진 그람음성세균, 세포벽을 가지지 않은 Mycoplasma 및 Thermoplasma군, 복잡하고 다양한 형태의 세포벽을 가지는 고세균으로 나눌 수 있다. 고세균의 표층구조는 펩티도글리칸 대신에 pseudopeptidoglycan 또는 당, 단백질, 당단백질, 이와는 다른 화학성분들로 이루어져 있다.
그람양성세균에서 펩티도글리칸은 약 20층으로 존재한다. 즉 그람양성세균의 세포벽의 80~90%가 펩티도글리칸으로 이루어져 있다. 일부 세균은 세포 주위에 펩티도글리칸의 단일층을 가지고 있지만, 대부분의 그람양성세균은 중첩된 여러겹(최대 25)의 펩티도글리칸층을 가지고 있다. 그람음성세균의 세포벽은 2~7nm정도 두께의 펩티도글리칸층과 그 바깥쪽을 둘러싸고 있는 7~8nm정도 두께의 외막으로 이루어져 있다. 그람양성세균의 경우 교차결합은 보통 펩티드 가교(peptide interbrige)의 형태로 이루어지는데 가교를 형성하는 아미노산의 종류와 수는 미생물에 따라 다르다. 실험에서 사용한 Staphylococcus aureus의 경우, 가교는 5개의 글리신으로 구성되어 있다.
그람음성세균에서 세포벽은 약 10%만이 펩티도글리칸으로 이루어져 있으며, 대장균과 몇몇 그람음성세균의 펩티도글리칸은 다 한층으로 존재한다. 그람음성세균에선 세포질막과 세포벽 사이에 주변세포질 공간(periplasmic space)이라 불리는 공간이 존재한다. Periplasmic space는 펩티도글리칸의 망상구조가 느슨하게 채워져 있고 여러 가지 기능을 가진 단백질들이 풍부하게 존재하기 때문에 젤(gel)상태에 가울것이라고 한다. 주변세포질 공간을 채우고 있는 물질을 주변세포질(periplasm)이라고 한다. 그람음성세균의 경우 교차결합은 한 글리칸 사슬의 DAP의 아미노기와 다음 글리칸 사슬의 말단 아미노산인 D-아랄닌의 카르복실기 사이의 펩티드 결합을 통해 이뤄진다.
고세균(Archaea)도 세포벽을 가지고 있기 때문에 그람양성 또는 음성으로 염색이 되지만 세포벽 구조와 화학성분은 세균과 상당히 다르다. 많은 그람양성 고세균은 그람양성세균과 유
<그림 4-1. 2번 배지를 전체적으로 봤을때>
분열하는 Staphylococcus aureus 세포의 투과전자현미경 사진을 보면 두꺼운 그람양성 세균의 세포벽을 볼 수 있다.
실험에서는 대장균과 황색포도상구균으로 그람양성균과 그람음성균을 구별하는 법을 배웠지만 이외의 다른 균으로는 그람양성세균은 Lactococcus lactis,Streptococcus 종, clostridium, Sarcina 종 등이 있으며 대부분 화학종속영양생물이 있고 그람음성세균으로는 시아노박테리아, 프로테어박테리아 등이 있다.
실험과정에서 몇 가지 주의할 것이 있다. 염색약은 묻으면 잘 지워지지 않으므로 장갑을 꼭 끼고 한다. 공기중에 노출되면 오염되니까 실험하는 곳을 물이나 에탄올로 세척한다. 백금이를 화염멸균하고 균을 취한다. 이때 두껍게 도말하면 관찰하기 어려우므로 백금이에 살짝 묻히듯 취한다. 또 불 근처에서 건조시킬때 빨리 건조시키기 위해 입으로 불거나 기구를 사용해서는 안된다. 침이나 먼지, 균이 들어가 오염되기 때문이다. 글라스 위에 물을 한방울 떨어뜨린 후 그 위에 균을 풀면서 지름 1~1.5cm 정도의 원모양으로 펼쳐 도말할 때 증류수는 퍼지게 하는 역할이다. 이것은 백금이로 편다. 증류수가 너무 많으면 잘 마르지 않아 그만큼 시간이 오래 걸리고 그만큼 공기 중의 다른 미생물이 실험 중인 균주에 들어가 오염될 가능성이 크다. 도말할 때 넓게 퍼지게 할수록 관찰하기 좋다. 도말 부위를 위로 한 상태에서 불꽃 중간에 슬라이드글라스를 3~4회 통과시켜서 표본을 열고정을 해야하는데 그 이유는 세척을 할 때, 세균이 씻겨 내려가는 것을 방지하기 위해서이다. 또 그람염색 중 뒷면에 증류수로 세척한 이유는 세균이 있는 앞면에 세척을 하면, 세균이 씻겨내려갈 수 있기 때문이다. 또 그람염색에서는 시약 순서대로 하는 것과 염색과 탈색 시간을 지키는 것이 아주 중요하다. 시간이 너무 짧으면 충분히 염색이 되지 않고, 또 오랫동안 염색하면 색이 너무 진해지며, 탈색시간인 10초를 넘으면, 그람양성균의 펩티도글리칸층이 아무리 두꺼워도 녹기 때문이다.
그람염색법은 미생물의 모양과 크기를 쉽게 관찰할 수 있다. Gram이 개발한 그람염색법에서 그람양성세균과 그람음성세균간의 차이점은 세포벽 구조의 차이에 기인한다. 그람음성세균의 세포벽은 다층 구조이며 아주 복잡하지만 그람양성세균의 세포벽은 단일층으로 되어 있고 더 두꺼우며 거의 모두 한 종류의 분자로 구성되어 있다. 그람양성세균과 그람음성세균에서 세포벽의 단단한 층(rigid layer)은 화학성분이 매우 유사한데 이를 펩티도글리칸이라 부른다. 그람양성세균의 세포벽은 세포질막 바쪽으로 20~80nm두께의 펩티도글리칸 단일층으로 이루어져 있다. 펩티도글리칸의 긴 사슬구조는 세포 주위를 둘러는 판을 형성하기 위하여 하나씩 연속적으로 생합성된다. 펩티도글리칸 사슬은 아미노산의 교차결합(cross-link)에 의하여 연결된다. 글리칸 사슬에서 당을 연결하는 배당결합(glycosidic bond)은 공유결합이지만 한 방향으로만 구조의 견고성을 줄 수 있다. 펩티도글리칸의 교차결합 후에만 X 및 Y방향으로 견고하게 된다. 교차결합은 세균의 종류에 라 그 정도가 다른데 교차결합이 많아질수록 더욱 견고해진다. 원핵세포는 표층구조의 차이에 따라 4군으로 나눌 수 있다. 즉 세포벽이 펩티도글리칸 단일층으로 이루어진 그람양성세균, 세포벽이 펩티도글리칸층과 그 바깥쪽을 둘러싸고 있는 외막(outer membrane)으로 이루어진 그람음성세균, 세포벽을 가지지 않은 Mycoplasma 및 Thermoplasma군, 복잡하고 다양한 형태의 세포벽을 가지는 고세균으로 나눌 수 있다. 고세균의 표층구조는 펩티도글리칸 대신에 pseudopeptidoglycan 또는 당, 단백질, 당단백질, 이와는 다른 화학성분들로 이루어져 있다.
그람양성세균에서 펩티도글리칸은 약 20층으로 존재한다. 즉 그람양성세균의 세포벽의 80~90%가 펩티도글리칸으로 이루어져 있다. 일부 세균은 세포 주위에 펩티도글리칸의 단일층을 가지고 있지만, 대부분의 그람양성세균은 중첩된 여러겹(최대 25)의 펩티도글리칸층을 가지고 있다. 그람음성세균의 세포벽은 2~7nm정도 두께의 펩티도글리칸층과 그 바깥쪽을 둘러싸고 있는 7~8nm정도 두께의 외막으로 이루어져 있다. 그람양성세균의 경우 교차결합은 보통 펩티드 가교(peptide interbrige)의 형태로 이루어지는데 가교를 형성하는 아미노산의 종류와 수는 미생물에 따라 다르다. 실험에서 사용한 Staphylococcus aureus의 경우, 가교는 5개의 글리신으로 구성되어 있다.
그람음성세균에서 세포벽은 약 10%만이 펩티도글리칸으로 이루어져 있으며, 대장균과 몇몇 그람음성세균의 펩티도글리칸은 다 한층으로 존재한다. 그람음성세균에선 세포질막과 세포벽 사이에 주변세포질 공간(periplasmic space)이라 불리는 공간이 존재한다. Periplasmic space는 펩티도글리칸의 망상구조가 느슨하게 채워져 있고 여러 가지 기능을 가진 단백질들이 풍부하게 존재하기 때문에 젤(gel)상태에 가울것이라고 한다. 주변세포질 공간을 채우고 있는 물질을 주변세포질(periplasm)이라고 한다. 그람음성세균의 경우 교차결합은 한 글리칸 사슬의 DAP의 아미노기와 다음 글리칸 사슬의 말단 아미노산인 D-아랄닌의 카르복실기 사이의 펩티드 결합을 통해 이뤄진다.
고세균(Archaea)도 세포벽을 가지고 있기 때문에 그람양성 또는 음성으로 염색이 되지만 세포벽 구조와 화학성분은 세균과 상당히 다르다. 많은 그람양성 고세균은 그람양성세균과 유
추천자료
- 가격2,000원
- 페이지수12페이지
- 등록일2014.01.21
- 저작시기2013.4
- 파일형식한글(hwp)
- 자료번호#902611

본 자료는 최근 2주간 다운받은 회원이 없습니다.


소개글